原型は白人至上主義者が作り出した似非学問
外見で人類集団を類別するなんて作業に勤しんでる連中に正義などあるわけないよね
おまけに反日至上主義者のアイヌ沖縄独立運動工作にも利用されてる始末
今時本気になってやってる奴は愚かで反道徳的なだけでしょう
3名無しさん@お腹いっぱい。
2021/03/23(火) 17:39:26.22ID:wWvMChJz 侮辱目的で大和民族を朝鮮民族と同一化するクソサヨク
いったい誰目線なの?とうに絶滅したジョウモン(笑)を持ち上げて国家転覆か?
15 名前:名無しさん@お腹いっぱい。[] 投稿日:2021/03/05(金) 12:24:27.11 ID:YnaVm7aJ [1/3]
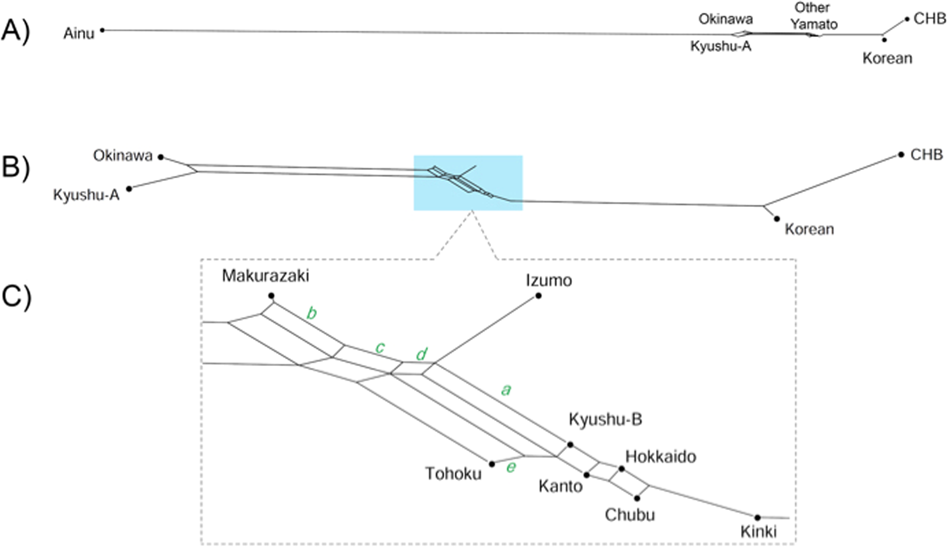
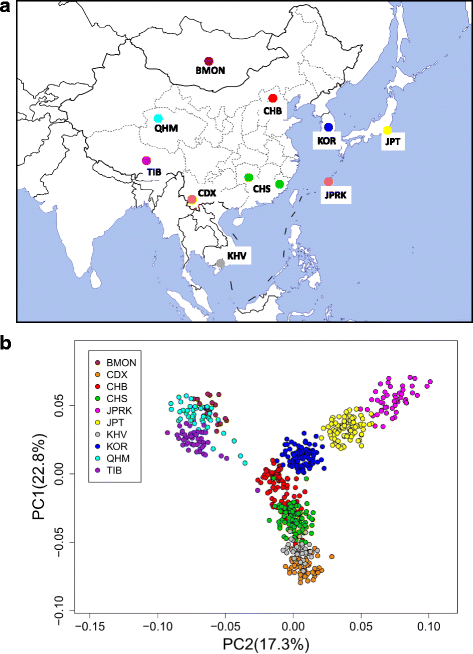
日本列島「本土」集団(ヤマト人)の「内部二重構造」モデルに関する研究(Jinam et al., 2021)
https://sicambre.at.webry.info/202102/article_4.html
近畿個体群がCHB・韓国人クラスタの最も近くに位置します。
2020年10月14日付けで『Journal of Human Genetics』に掲載された報告では、朝鮮半島に地理的に近い九州北部より、四国・近畿地方に渡来人の遺伝子が強く見られたと述べられています。
https://www.asahi.com/articles/ASNBG3HLFNBFULBJ00P.html

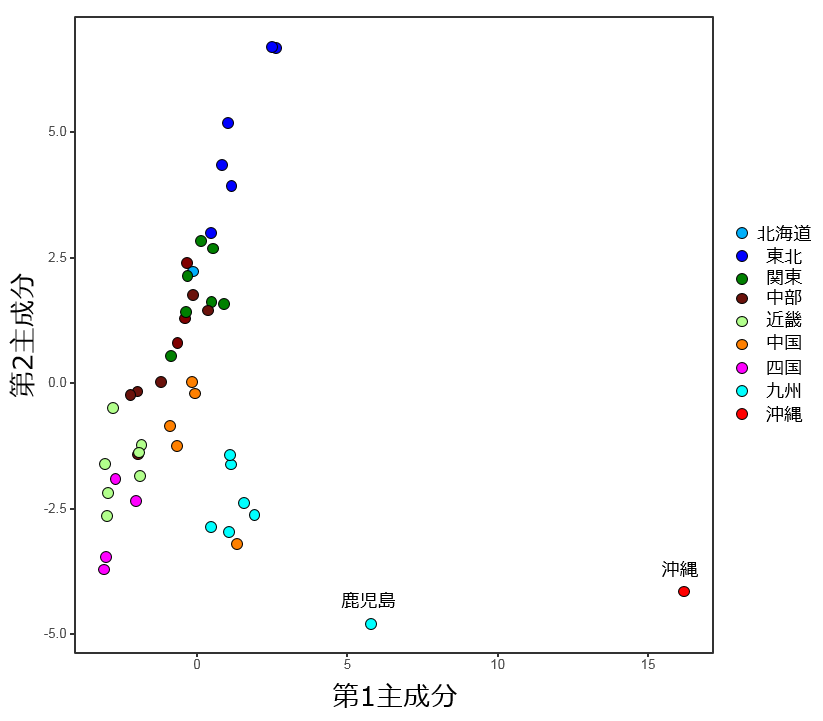
中国朝鮮-近畿四国-中国-中部-関東北海道-東北
愛媛の歌人正岡子規「人間としては下等の地に居る、死に歌詠みの公家たち」


赤色→北海道、沖縄、北東北、鹿児島
橙色→南東北、関東、新潟、九州
黄色→近畿、瀬戸内、朝鮮半島
(国立科学博物館)
>関東出土の縄文人骨の多くは頑丈で、西日本出土の人骨は小さい(小片保)
>東日本では動物性食料資源の捕獲や調理のための道具が発達し、西日本では植物性食料資源を利用するための道具が多い(赤沢威)
http://ww3.ctt.ne.jp/~seijiham/seikatu/kyoiku/shotou/tai/taii_m_prf.gif
【身長(5~17歳、文科省「学校保健統計調査」2018年)】
東北・北海道>北陸>首都圏>北関東>関西>九州四国>東海>中国
【体重(5~17歳、文科省「学校保健統計調査」2018年)】
東北・北海道>北関東>北陸>九州四国>首都圏>関西>東海>中国
いったい誰目線なの?とうに絶滅したジョウモン(笑)を持ち上げて国家転覆か?
15 名前:名無しさん@お腹いっぱい。[] 投稿日:2021/03/05(金) 12:24:27.11 ID:YnaVm7aJ [1/3]
日本列島「本土」集団(ヤマト人)の「内部二重構造」モデルに関する研究(Jinam et al., 2021)
https://sicambre.at.webry.info/202102/article_4.html
近畿個体群がCHB・韓国人クラスタの最も近くに位置します。
2020年10月14日付けで『Journal of Human Genetics』に掲載された報告では、朝鮮半島に地理的に近い九州北部より、四国・近畿地方に渡来人の遺伝子が強く見られたと述べられています。
https://www.asahi.com/articles/ASNBG3HLFNBFULBJ00P.html
中国朝鮮-近畿四国-中国-中部-関東北海道-東北
愛媛の歌人正岡子規「人間としては下等の地に居る、死に歌詠みの公家たち」
赤色→北海道、沖縄、北東北、鹿児島
橙色→南東北、関東、新潟、九州
黄色→近畿、瀬戸内、朝鮮半島
(国立科学博物館)
>関東出土の縄文人骨の多くは頑丈で、西日本出土の人骨は小さい(小片保)
>東日本では動物性食料資源の捕獲や調理のための道具が発達し、西日本では植物性食料資源を利用するための道具が多い(赤沢威)
http://ww3.ctt.ne.jp/~seijiham/seikatu/kyoiku/shotou/tai/taii_m_prf.gif
【身長(5~17歳、文科省「学校保健統計調査」2018年)】
東北・北海道>北陸>首都圏>北関東>関西>九州四国>東海>中国
【体重(5~17歳、文科省「学校保健統計調査」2018年)】
東北・北海道>北関東>北陸>九州四国>首都圏>関西>東海>中国